|
Virgin and Child, Enthroned, The Master of Moulins, 1499 |
|
|
|
Virgin and Child, Enthroned, The Master of Moulins, 1499 |
|
|
Science, Vol 300, Issue 5617, 297 , 11 April 2003
Molecular Correlates of Primate Nuclear Transfer Failures
Calvin
Simerly,1* Tanja Dominko,1*![]() Christopher Navara,1 Christopher Payne,1 Saverio Capuano,1
Gabriella Gosman,1 Kowit-Yu Chong,1 Diana Takahashi,1
Crista Chace,1 Duane Compton,2 Laura Hewitson,1
Gerald Schatten1
Christopher Navara,1 Christopher Payne,1 Saverio Capuano,1
Gabriella Gosman,1 Kowit-Yu Chong,1 Diana Takahashi,1
Crista Chace,1 Duane Compton,2 Laura Hewitson,1
Gerald Schatten1![]()
1
Pittsburgh Development Center, Magee-Womens Research Institute, Departments of
Obstetrics-Gynecology-Reproductive Sciences & Cell Biology-Physiology,
University of Pittsburgh School of Medicine, Pittsburgh, PA 15213, USA.
2 Department of Biochemistry, Dartmouth Medical School, Hanover, NH
03755, USA.
* These authors contributed equally to this
work
![]() Present address: CellThera, Worcester, MA 01605.
Present address: CellThera, Worcester, MA 01605.
![]() To whom correspondence should be addressed. E-mail:
gschatten@magee.edu
To whom correspondence should be addressed. E-mail:
gschatten@magee.edu
Somatic cell nuclear transfer (SCNT) (1) in nonhuman primates could accelerate medical research by contributing identical animals for research and clarifying embryonic stem cell potentials (2). Although rhesus embryos begin development after embryonic cell nuclear transfer (ECNT) (3-5), there has only been one report of rhesus births after ECNT (3), and that report has not been replicated.
Here, molecular obstacles were identified using 716 rhesus oocytes in four experimental studies: set A, SCNT [rhesus cumulus, umbilical cord blood, epithelial-derived fibroblasts, and inner cell mass-derived precursor embryonic stem cells; 193 oocytes; 62.8% nuclear transfer (NT) success assayed by interphase nucleus formation], and set B, ECNT from dissociated 16- to 32-cell stage embryos (381 oocytes; 97.2% NT success), because ECNT success is greater than SCNT (1). Because meiotic spindle removal appeared to be responsible for these NT failures, we performed two additional experiments in which either we did not remove the spindle (set C) or we removed and reinserted it (set D). In set C, NTs into concurrently fertilized oocytes generated tetraploids (55 oocytes; 54.4% success), whereas in set D, fertilization of reconstituted oocytes (that had previously been enucleated and then renucleated) generated diploids (95 oocytes; 67.1% success).
Rhesus NTs (6) look superficially normal, yet no pregnancies resulted from 33 embryos transferred into 16 surrogates (compared with seasonably variable 28 to 66% pregnancy rates by assisted reproduction) (7). DNA and microtubule imaging showed disarrayed mitotic spindles with misaligned chromosomes (Fig. 1A; all 116 ECNTs and all 30 SCNTs examined displayed aberrant spindles). Despite these defects, cleavages continue, but unequal chromosome segregations produce aneuploid embryos.

Fig.
1. Faulty mitotic spindles produce aneuploid embryos after
primate
nuclear
transfer.
(A) Defective NT mitotic spindle with misaligned chromosomes. Centrosomal
NuMA at meiosis (B) and mitosis (C), but not in mitotic spindles
after NT (D). The centrosomal kinesin HSET is also missing after NT (E),
but not centromeric Eg5. (F). Bipolar mitotic spindles with aligned
chromosomes and centrosomal NuMA after NT into fertilized eggs (G). DNA,
microtubule, NuMA, and kinesin imaging as in (7,
8). Blue, DNA; red, ![]() -tubulin;
green, NuMA in (B), (C), (D), and (G); HSET in (E); and Eg5 in (F). Scale bar,
10 µm.
[View Larger Version of this Image (42K GIF file)]
-tubulin;
green, NuMA in (B), (C), (D), and (G); HSET in (E); and Eg5 in (F). Scale bar,
10 µm.
[View Larger Version of this Image (42K GIF file)]
NuMA (Nuclear-Mitotic Apparatus), a matrix protein responsible for spindle pole assembly (8), concentrates at centrosomes in unfertilized meiotic (Fig. 1B) and fertilized mitotic cells (Fig. 1C). After NT, NuMA is not detected on the abnormal mitotic spindles (Fig. 1D) or in enucleated oocytes. HSET and Eg5 are mitotic kinesin motors (8, 9). HSET, found during meiosis and mitosis, is not detected in NT spindles (Fig. 1E). Eg5 detects centromere pairs at meiosis and mitosis, including misaligned ones on NT spindles (Fig. 1F). Thus, meiotic spindle removal depletes the ooplasm of NuMA and HSET, both vital for mitotic spindle pole formation.
Normal spindles found in tetraploids suggest meiotic spindle removal as the source of NT anomalies. In tetraploids, chromosomes aligned properly on bipolar spindles with centrosomal NuMA (Fig. 1G). NT mitotic spindles could be distinguished from the fertilized spindle by the sperm tail. Similarly, fertilization of reconstituted oocytes resulted in apparently normal divisions. Thus, manipulation of the embryos alone was not the cause of the problem, and proper mitotic spindles can be organized around somatic chromosomes if the meiotic spindle is left intact.
Primate NT appears to be challenged by stricter molecular requirements for mitotic spindle assembly than in other mammals. In cattle, the somatic centrosome is transferred during NT (10), whereas mice rely on the oocyte's maternal centrosome (11). Also, NuMA and HSET are not exclusively concentrated on the meiotic spindle in mammals other than primates (8). With current approaches, NT to produce embryonic stem cells in nonhuman primates may prove difficult--and reproductive cloning unachievable.
REFERENCES AND NOTES
1. I. Wilmut, Nature Med. 8, 215 (2002) [CrossRef][ISI][Medline].
2. J. A. Thomson, et al., Science 282, 1145 (1998) [Abstract/Free Full Text].
3. L. Meng, J. J. Ely, R. L. Stouffer, D. P. Wolf, Biol. Reprod. 57, 454 (1997) [Abstract].
4. T. Dominko, et al., Cloning 1, 143 (1999) [CrossRef].
5. S. M. Mitalipov, R. R. Yeoman, K. D. Nusser, D. P. Wolf, Biol. Reprod. 66, 1367 (2002) [Abstract/Free Full Text].
6. Materials and methods are available as supporting material on Science Online.
7. L. Hewitson, et al., Nature Med. 5, 431 (1999) [CrossRef][ISI][Medline].
8. V. Mountain, et al., J. Cell Biol. 147, 351 (1999) [Abstract/Free Full Text].
9. A. Blangy, et al., Cell 83, 1159 (1995) [ISI][Medline].
10. C. S. Navara, N. L. First, G. Schatten, Dev. Biol. 162, 29 (1994) [CrossRef][ISI][Medline].
11. G. Schatten, Dev. Biol. 165, 299 (1994) [CrossRef][ISI][Medline].
12. We thank S. Doxsey, J. Jones, O. Khorram, A. Lewis, V. Mountain, T. Plant, J. St. John, and T. Stearns and Serono Reproductive Biology Institute (Rockland, MA 02370) for hormones and we gratefully acknowledge NIH support.
10.1126/science.1082091
Include this information when citing this paper.
____________________________________________________________
Science,
Vol 301, Issue 5639, 1482 ,
12 September 2003
[DOI: 10.1126/science.1085871]
Comment on "Molecular Correlates of Primate Nuclear Transfer Failures"
We believe that Simerly et al. (1) have reported important new data but have overstated their conclusions. Their preliminary observations highlight the importance of egg quality—centrosome and spindle competence, specifically—in nuclear transfer (NT) procedures. However, the data are scant and their interpretation is compromised by the multiplicity of methods used. In our own experience, multiple factors including donor and recipient cell cycle stage and oocyte age can influence the integrity of the spindle complex. Simerly et al. buttressed their claim that primates are unique in the specific absence of NuMA (nuclear-mitotic apparatus protein) or HSET based only on immunofluorescence. However, one cannot conclude from these data that the oocyte was depleted of these proteins—only that they were not detected where expected using the techniques described. The occurrence of abnormal spindles has long plagued NT in most species, and few comparable immunocytochemical studies have been reported to support the conclusion that failed spindle complexes in other species would not lack those markers as well. More comprehensive investigations of NT-induced centrosome-spindle perturbations will reveal the true impact of these manipulations on the efficiency of NT on a species-byspecies basis.
Simerly
et al. transferred only 33 rhesus embryos into 16 surrogates
and concluded that reproductive cloning in primates may be unachievable.
In a related news article (2),
the senior author of (1)
stated that it is almost as if someone "drew a sharp line between
old-world primates—including people—and other animals, saying,
`I'll let you clone cattle, mice, sheep, even rabbits and cats,
but monkeys and humans require something more.'" In our own hands, it
took dozens of embryos to generate Dolly (3),
more than 150 embryos to generate the first cloned mouse pup (4),
and 586 embryos to establish the first two pregnancies in pigs
(5).
Hundreds of other studies have ended with no pregnancies at all. We
suggest that given the potential importance of nuclear transfer
techniques in human cell therapies, conclusions regarding the
efficiency of human NT extrapolated from animal data should be
tempered with abundant caution. Robert Lanza
Young Chung
Michael D. West
Advanced Cell Technology
One Innovation Drive
Worcester, MA 01605, USA
E-mail:
rlanza@advancedcell.com
Keith H.
S. Campbell
Division of
Biological
Sciences
University of Nottingham
Loughborough, UK
|
|
References
1. C. Simerly et al., Science 300, 297 (2003).[Free Full Text]
2. G. Vogel, Science 300, 225 (2003).[Abstract/Free Full Text]
3. I. Wilmut, A. E. Schnleke, J. McWhir, A. J. Kind, K. H. S. Campbell, Nature 385, 810 (1997).[CrossRef][ISI][Medline]
4. Y. Chung, unpublished data.
5. I. A. Polejaeva et al., Nature 407, 86 (2000).[CrossRef][ISI][Medline]
17 April 2003; accepted 31 July 2003
________________________________________________________________
Science,
Vol 301, Issue 5639, 1482 , 12 September 2003
[DOI: 10.1126/science.1087756]
Response to Comment on "Molecular Correlates of Primate Nuclear Transfer Failures"
Space constraints precluded publication of bovine nuclear transfer (NT) data (Fig. 1) that may allay the concerns of Lanza et al (1). After primate NT, disorganized spindles with chaotically distributed chromosomes are always observed (2). Bovine NTs, however, organize a single microtubule aster emanating from the juxtanuclear NT centrosome at first interphase (Fig. 1A). Mitotic bovine NTs frequently assemble symmetrical bipolar spindles with precisely aligned chromosomes [Fig. 1B, blue and (2)], a situation never observed after primate NT. These and centrosome and kinesin NT data are subjects of a forthcoming report (3).
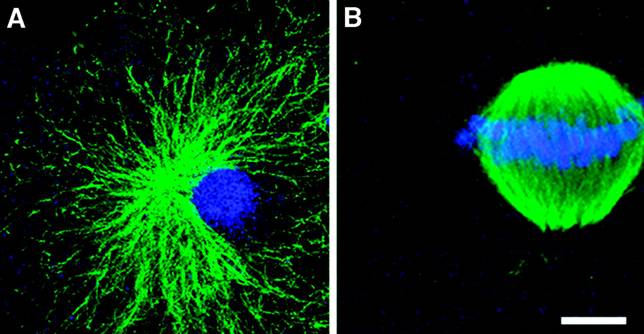
Fig. 1. Chromosomes align on symmetrical mitotic spindles after bovine nuclear transfer, unlike the chaotic spindles detected after primate NT (4). (A) Somatic cell nuclear transfer in cattle oocytes results in a single radially arrayed microtubule aster at first interphase. (B) At first mitosis, bovine NTs organize accurate bipolar mitotic apparatus (green) with aligned chromosomes (blue). Scale bar: A, 10 µm; B, 5 µm.
Without benefit of this additional knowledge, Lanza et al. fairly raise the uncertainty in interpreting NT results, especially in extrapolating assisted reproductive technology (ART) successes or failures between species. For example, >90% fertilization is routinely achieved using intracytoplasmic sperm injection (ICSI) in both humans and nonhuman primates (resulting in the global adoption of ICSI by ART clinics), whereas ICSI has produced few mice or cattle (4). Because NT successes vary, comparisons between Dolly's birth after 29 embryo transfers into 13 ewes (5) and our lack of established pregnancies after 33 rhesus embryo transfers are not reasonable. Primate ART, including NT, is uniquely challenging (6): Oocyte availability and quality, synchronized surrogates, cleavage-stage embryo transfers versus blastocysts transfers, and reproductive seasonality all present practical limitations. And unlike mice and domestic mammals, primates carry singletons. Finally, while the protocols for mouse and bovine NT are species-specific, primate laboratories must extrapolate from human ART clinics for simian-specific optimizations.
Although human embryonic stem cells (hESCs) derived after NT might overcome immune rejection, NT epigenetic abnormalities (7) and microchimerism in transplantation patients (8) encourage alternate strategies for tolerance or managing incompatibilities. In light of the available evidence, we maintain our original conclusion (2) that "[w]ith current approaches, [primate] NT to produce embryonic stem cells may prove difficult—and reproductive cloning unachievable." We agree with Lanza et al. (1) that interpretations regarding both NT and hESC data in primates, human (9, 10) and nonhuman (5, 11) alike, should be "tempered with abundant caution."
G. Schatten
C. Navara
C. Payne
S. Capuano
G. Gosman
K.-Y. Chong
D. Takahashi
C. Chace
L. Hewitson
C. Simerly
Pittsburgh Development Center,
Magee-Womens Research Institute,
Department of Obstetrics-Gynecology-
Reproductive Sciences and Cell
Biology-Physiology,
University of Pittsburgh School of Medicine
Pittsburgh, PA 15213, USA
E-mail:
gschatten@magee.edu
D. Compton
Department of Biochemistry
Dartmouth Medical School
Hanover, NH 03755, USA
T. Dominko
CellThera
Worcester, MA 01605, USA
References
1. R. Lanza, Y. Chung, M. D. West, K. H. S. Campbell, Science 301, 1482 (2003); www.sciencemag.org/cgi/content/full/301/5639/1482b.
2. C. Simerly et al., Science 300, 297 (2003).[Free Full Text]
3. C. Navara et al., unpublished results (2003).
4. G. Schatten, L. Hewitson, C. Simerly, P. Sutovsky, G. Huszar, J. Law Med. Ethics 26, 29 (1998).[ISI][Medline]
5. I. Wilmut, A. E. Schnleke, J. McWhir, A. J. Kind, K. H. S. Campbell, Nature 385, 810 (1997).[CrossRef][ISI][Medline]
6. L. Hewitson et al., Nature Med. 5, 431 (1999).[CrossRef][ISI][Medline]
7. K. Hochedlinger, R. Jaenisch, N. Engl. J. Med. 349, 275 (2003).[Free Full Text]
8. T. E. Starzl et al., Lancet 361, 1502 (2003).[CrossRef][ISI][Medline]
9. J. B. Cibelli et al., E-biomed J. Regener. Med. 2, 25 (2001).[CrossRef]
10. R. P. Lanza, J. B. Cibelli, M. D. West, Nature Biotechnol. 20, 117 (2002).[CrossRef][ISI][Medline]
11. J. B. Cibelli et al., Science 295, 819 (2002).[Free Full Text]
9
June 2003; accepted 8 August 2003
10.1126/science.1087756
Include this information when citing this paper.

