|
Virgin and Child, Enthroned, The Master of Moulins, 1499 |
ADULT
STEM
CELLS
FOR TISSUE
REPAIR
— A
NEW
THERAPEUTIC
CONCEPT?
|
|
|
Virgin and Child, Enthroned, The Master of Moulins, 1499 |
ADULT
STEM
CELLS
FOR TISSUE
REPAIR
— A
NEW
THERAPEUTIC
CONCEPT?
|
|
The New England
Journal of Medicine.
Volume 349:570-582, August 7,
2003,Number 6
Adult human stem cells that are intrinsic to various tissues have been described and characterized, some of them only recently. These cells are capable of maintaining, generating, and replacing terminally differentiated cells within their own specific tissue as a consequence of physiologic cell turnover or tissue damage due to injury.1 Hematopoietic stem cells that give rise to blood cells and move between bone marrow and peripheral blood are the best-characterized adult stem cells in humans. Recent data suggest that adult stem cells generate differentiated cells beyond their own tissue boundaries, a process termed ‘developmental plasticity.’ In this review we focus on in vivo models of adult stem cells derived from bone marrow and peripheral blood and their potential therapeutic applications.
|
ADULT STEM CELLS AND THEIR POTENTIAL FOR DEVELOPMENTAL PLASTICITY |
Stem cells are defined as cells that have clonogenic and self-renewing capabilities and that differentiate into multiple cell lineages.2 Whereas embryonic stem cells are derived from mammalian embryos in the blastocyst stage and have the ability to generate any terminally differentiated cell in the body, adult stem cells are part of tissue-specific cells of the postnatal organism into which they are committed to differentiate. Phenotypically characterized adult stem cells are listed in Table 1.1,3
|
|
Table 1.
and Primary
Direction
|
The hematopoietic system has traditionally been seen as an organized, hierarchic system with multipotent, self-renewing stem cells at the top, committed progenitor cells in the middle, and lineage-restricted precursor cells, which give rise to terminally differentiated cells, at the bottom.2 However, this classic paradigm of stem-cell differentiation restricted to its organ-specific lineage is being challenged by the suggestion that adult stem cells, including hematopoietic stem cells, retain a previously unrecognized degree of developmental plasticity that allows them to differentiate across boundaries of lineage, tissue, and germ layer (Figure 1). The hierarchical view no longer seems correct.3,4
|
|
|
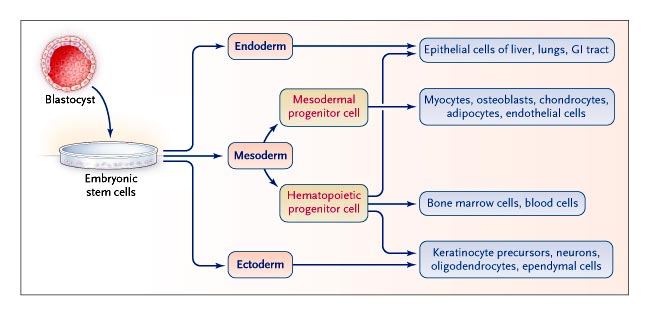
Figure 1. Model of Embryonic and Adult Stem-Cell Differentiation along and across Germ-Layer Boundaries. Embryonic stem cells differentiate into three germ-layer–type cells when cultured under appropriate conditions. Some data suggest that postnatal mesodermal hematopoietic progenitor cells destined to differentiate along their own pathway may, like embryonic stem cells, cross germ-layer boundaries to generate endodermal or ectodermal tissue cells. GI denotes gastrointestinal. |
The molecular mechanisms of lineage switches within the hematopoietic system have been studied extensively,5 but the mechanisms that determine transitions in the fate of adult stem cells remain poorly understood. The results of recent studies of the plasticity of adult stem cells, which contradict the dogma that the differentiation and commitment of adult stem cells are restricted to their own tissue, are the subject of intense discussion. These findings demand the most stringent criteria for providing conclusive evidence. To prove that stem cells derived from bone marrow and peripheral blood, including hematopoietic stem cells, are indeed transformed into solid-organ–specific cells, several conditions must be met. First, the origin of the exogenous cell integrated into solid-organ tissue must be documented by cell marking, preferably at the single-cell level. Cells also should be processed with a minimum of ex vivo manipulation (e.g., culturing), which may make them more susceptible to crossing lineages. Second, the exogenous cell must be shown to have become an integral morphologic part of the newly acquired tissue. Third, and most important, the transformed cell must be shown to have acquired the function of the particular organ into which it has been integrated, both by expressing organ-specific proteins and by showing specific organ function.
|
PRECLINICAL IN VIVO STUDIES |
Various stem-cell preparations originating from hematopoietic tissue have been used in in vivo studies of stem-cell plasticity, including unselected cells from bone marrow, purified hematopoietic stem cells, and single hematopoietic stem cells that originate from either bone marrow or peripheral blood. Because hematopoietic tissue (from both bone marrow and peripheral blood) harbors a population of heterogeneous stem cells that includes hematopoietic stem cells, mesenchymal stem cells, multipotent adult progenitor cells, and endothelial precursor cells, some of the contradictory findings that have been reported may be explained by the use of different sources and preparations of stem cells.
|
ADULT STEM CELLS DERIVED FROM BONE MARROW |
Initial in vivo studies were performed with the use of marked, unselected cells derived from bone marrow. After transplantation of the cells into animals that had undergone a conditioning treatment, there was evidence that the cells formed nonlymphohematopoietic tissue, such as muscle fibers,6 hepatocytes,7 microglia and astroglia,8 and neuronal tissue.9,10
Although the differentiation of stem cells derived from bone marrow into organ-specific, nonlymphohematopoietic cells has been described, the origin of stem cells responsible for the formation of nonlymphohematopoietic tissue remains obscure. Thus, to prove that stem cells from bone marrow — and, in particular, hematopoietic stem cells — are capable of forming solid-organ tissue cells, purified stem cells with a specific marker had to be transplanted and shown to generate specific functional tissue cells.11
Donor-derived muscle fibers that express dystrophin have been identified after sex-mismatched transplantation of purified and phenotypically characterized hematopoietic stem cells.12 Lagasse and colleagues13 went one step further by providing proof of functionality: in an animal model of tyrosinemia type I, transplantation of as few as 50 purified hematopoietic stem cells restored both the hematopoietic and biochemical liver functions in the recipient by correcting the genetic aberration. In a similar fashion, hepatocytes that express human albumin were identified in immunodeficient mice in which purified human hematopoietic stem cells had been transplanted.14 In other studies, purified stem cells, including hematopoietic stem cells, have been shown to generate functioning cardiomyocytes and vascular structures,15,16 as well as neointimal smooth-muscle cells and endothelial cells that contribute to arterial remodeling in various models of vascular lesions.17
Further in vivo evidence that hematopoietic stem cells are capable of multifunctional differentiation was provided by Krause and colleagues.18 Using a limiting-dilution technique, they showed that single cells differentiated into mature hematopoietic cells and into mature epithelial cells of the skin, lungs, and gastrointestinal tract. However, because of the localization of the purported transdifferentiated progeny and the lack of evidence of organ-specific function of those cells, the physiologic relevance of the reported observations is not clear. In similar experimental settings, transplantation of a single hematopoietic stem cell or cultured cells derived from a single hematopoietic stem cell resulted not only in hematopoietic engraftment but also in retinal neovascularization19 [creation of new blood vessel] and the generation of functioning glomerular mesangial cells [kidney tissue].20
|
ADULT STEM CELLS DERIVED FROM PERIPHERAL BLOOD |
Since progenitor cells derived from bone marrow can reach the target solid organ through the peripheral blood,21,22 determining whether peripheral-blood stem cells follow a differentiation pathway specific to solid organs, similar to that seen for stem cells derived from bone marrow, was the logical next step.
Several investigators have reported that circulating human stem cells, mobilized into the peripheral blood by cytokine administration, contribute to the generation of nonlymphohematopoietic tissue. Endothelial progenitor cells mobilized by recombinant human granulocyte–macrophage colony-stimulating factor and recombinant human granulocyte colony-stimulating factor were shown to contribute to ocular neovascularization in mice23 and neovascularization of ischemic myocardium in rats,24 respectively. Orlic and coworkers25 provided evidence of the generation of cardiomyocytes in a murine model of myocardial infarction after a cytokine-induced increase in the concentration of circulating stem cells. These changes resulted in improved ventricular function and survival.25 However, in a different experimental setting involving a nonhuman primate model of myocardial infarction and reperfusion, cytokine-induced mobilization of stem cells did not result in a favorable outcome.26
Clinical in Vivo Studies
|
ADULT STEM CELLS DERIVED FROM BONE MARROW AND PERIPHERAL BLOOD |
Because the experimental conditions in a preclinical setting are currently far more sophisticated than those available in a clinical setting, the mechanisms underlying the reported clinical observations should be interpreted with caution. The same is true when extrapolating knowledge about stem-cell function in nonprimate species to humans.
The ability of human progenitor cells derived from bone marrow to generate nonlymphohematopoietic tissue has been studied in allogeneic sex-mismatched transplants. The initial clinical studies were performed by Horwitz and colleagues,27 who claimed that stem cells derived from bone marrow led to improved osteogenesis [creation of new bone] in children with osteogenesis imperfecta. Subsequently, two groups independently reported the presence of donor cells that were positive for the Y chromosome in liver tissue after male-into-female bone marrow transplantation or female-into-male liver transplantation. Theise and colleagues28 reported the presence of donor-derived hepatocytes in liver tissue at levels ranging from 4 to 43 percent and cholangiocytes at levels ranging from 4 to 38 percent. At the same time, Alison et al.29 also reported donor-derived hepatocytes, although at lower levels (0.5 to 2 percent).
In another study of patients who underwent liver transplantation, Kleeberger et al.30 confirmed liver-tissue chimerism by genotyping cells that had been microdissected with a laser and immunolabeled with cytokeratin, using DNA analysis of highly polymorphic short tandem repeats. Cholangiocyte chimerism was an almost universal finding in the early period after transplantation, whereas hepatocyte chimerism tended to occur later, coinciding with recurrent hepatitis. Epithelial cells of the esophagus, stomach, small intestine, and colon of donor origin were reported at a frequency of 0.4 to 4.6 percent in patients who underwent sex-mismatched bone marrow transplantation in whom graft-versus-host disease or ulcer formation subsequently developed.31 Translating their observations in rodents to a clinical level, Mezey et al.32 identified rare clusters of donor-derived neuronal cells containing the Y chromosome at a frequency of up to 7 per 10,000 neurons in the hippocampus and cerebral cortex of recipients of bone marrow transplants. In a similar transplantation setting, donor-derived Purkinje neurons were identified in the brains of adults who had received bone marrow transplants.33
Indications that circulating stem cells can contribute to the formation of solid-organ tissue derive from studies of solid-organ transplantation. Quaini et al.34 and Müller et al.35 have reported male chimerism in heart allografts from female donors. In addition to the effect of direct migration of cardiomyocytes from adjacent recipient tissue into the allograft, the researchers postulate that circulating stem cells from the transplant recipient contribute to ventricular remodeling in heart allografts. Similarly, Lagaaij et al.36 have documented endothelial-cell chimerism in patients with rejection of renal allografts. Circulating donor-derived endothelial cells have been identified in the peripheral blood of recipients of stem-cell transplants by determination of their origin either from transplanted circulating angioblasts or from transplanted stem cells that differentiated in vivo into endothelial cells.37
We and our colleagues recently reported the presence of XY-positive hepatocytes and epithelial cells in five female recipients of peripheral-blood stem-cell allografts mobilized by recombinant human granulocyte colony-stimulating factor from male donors.38 Donor-derived, XY-positive, nonlymphohematopoietic cells were identified at frequencies ranging from 0 to 7 percent in the skin, gut, and liver of all five female stem-cell recipients as early as day 13 and up to day 354 after transplantation with allogeneic stem cells. Although two conditions for proof that donor-derived male peripheral-blood cells contributed to the formation of solid-organ tissue in female recipients were met with the use of the Y-chromosome marker and morphologic evidence of donor cells integrated into the recipient's solid-organ tissue, we did not demonstrate that donor-derived cells, whether located within mature or immature cell compartments, expressed functions specific to solid-organ tissue.
Donor-derived keratinocytes that were positive for the Y chromosome and cytokeratin were found at a frequency ranging from 3.7 to 14.8 percent in six patients who underwent transplantation of sex-mismatched peripheral-blood stem cells.39 However, when the same epidermal skin cells were cultured over a period of 18 to 32 days through multiple passages to eliminate any contaminating lymphohematopoietic cells, Y-chromosome–specific sequences from the donor-derived cells could not be detected. One might speculate that the donor-derived cells found in skin tissue were artifacts of the engulfment of the recipient's skin cells by donor macrophages; that cells bearing the Y-chromosome–specific sequences were lost after several passages in culture; or that the donor cells had little or no proliferative capacity in vitro.39 Also, donor-derived buccal epithelial cells were identified in recipients of sex-mismatched peripheral-blood stem-cell transplants up to 4.5 years after transplantation at a frequency between 0.8 percent and 12.7 percent.40 Given the number and variety of clinical studies that have already been performed (Table 2), and the lack of data from studies of large animals — in particular, primates — caution should be exercised in predicting a potential clinical benefit.
|
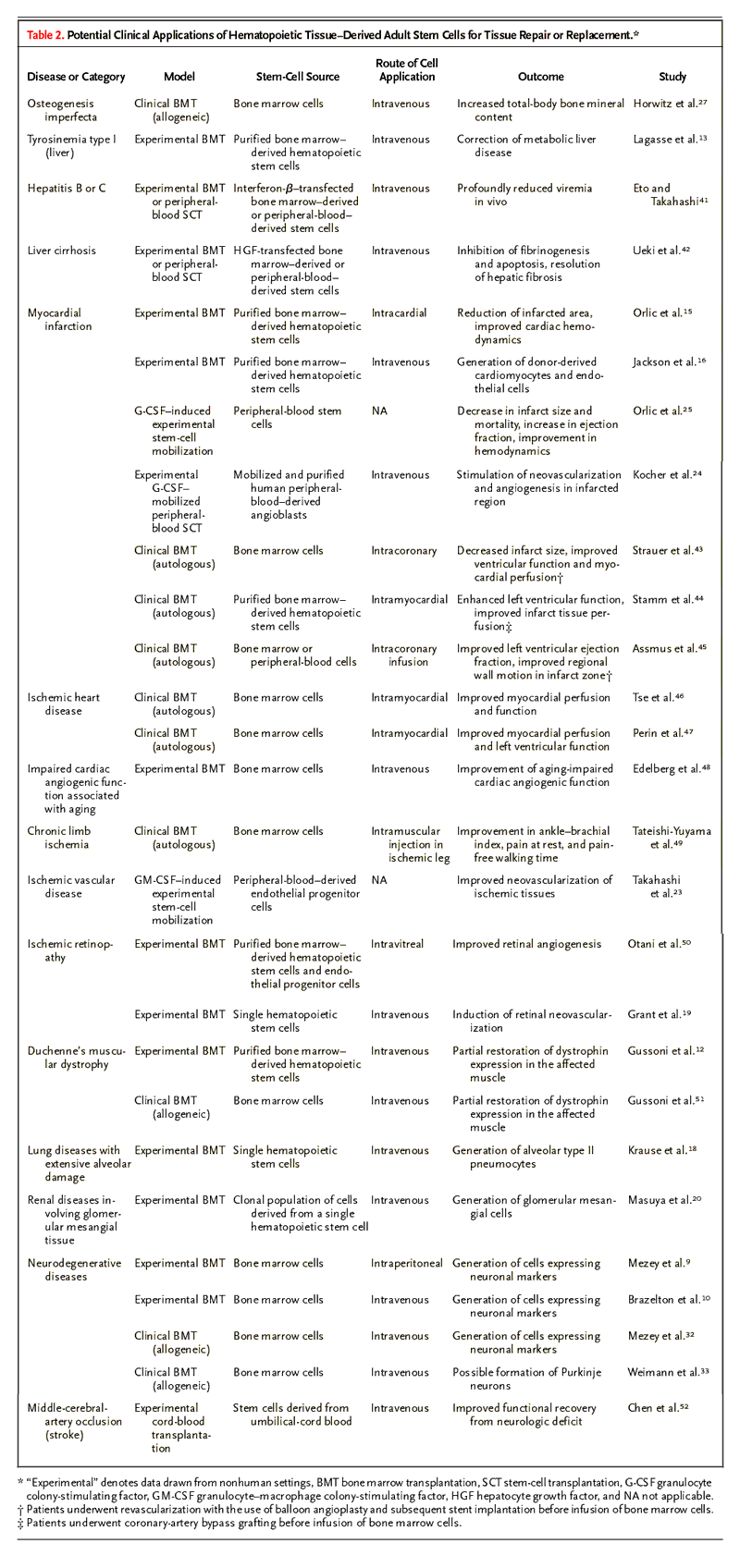
Table 2. Potential Clinical Applications of Hematopoietic Tissue–Derived Adult Stem Cells for Tissue Repair or Replacement |
|
|
|
LIMITATIONS IN IDENTIFYING CHIMERIC CELLS |
The interpretation of the data on tissue chimerism could be hindered by technical conditions. Y-chromosome–positive nuclei must be unequivocally identified as belonging to nonlymphohematopoietic solid-organ–specific cells integrated into female tissue, in order to rule out the possibility that donor-derived inflammatory cells, such as infiltrating lymphocytes or macrophages, are mistakenly identified as solid-organ–specific cells.53 One way to demonstrate that Y-chromosome–positive nuclei are truly associated with solid-organ–specific cells is to perform concomitant or sequential staining, including fluorescence in situ hybridization staining, on the same thin tissue section instead of staining serial sections separately.38
|
REPORTS THAT QUESTION DEVELOPMENTAL PLASTICITY |
Two reports have been published that question the potential of cells derived from hematopoietic tissue to undergo cell fate transition and in so doing contribute to cell formation in solid-organ tissue. Castro et al.54 failed to detect neural-like cells in the brains of mice that received purified stem cells derived from bone marrow or unfractionated bone marrow cells. Similarly, after transplanting single hematopoietic stem cells into conditioned recipient mice, Wagers et al.55 found very few donor-derived cells in nonlymphohematopoietic tissue. As discussed in detail elsewhere with regard to the findings of Castro et al. and Wagers et al., the interpretation of negative results depends on the experimental system in which the hypothesis is tested and on its specificity and sensitivity.56,57,58
|
CELL FUSION |
Recently, the validity of stem-cell plasticity as a mechanism for generating nonlymphohematopoietic tissue has been questioned. Some have suggested that previously overlooked cell fusion may explain the findings in the studies described above.
Confirming earlier studies performed under rather nonphysiologic culture conditions,59,60 two in vivo studies of mice with a fatal metabolic liver disease have provided evidence that bone marrow cells from normal donors generate healthy hepatocytes by forming hybrid cells that contain both donor and host genes.61,62 Most of the fused cells had tetraploid or hexaploid DNA content. In contrast, cytogenetic analysis of bone marrow–derived cells specific to solid organs in preclinical allogeneic transplant studies revealed a diploid karyotype.20,63 Donor-derived, solid-organ–specific cells in patients who have received allogeneic bone marrow or peripheral-blood stem-cell transplants have also been identified as diploid,28,29,38,39,64 except in liver tissue, where polyploidy is not uncommon.
Furthermore, numerous cytogenetic analyses of bone marrow–biopsy specimens from patients undergoing allogeneic stem-cell transplantation have been characterized by euploidy, except in diseased tissue. It is possible that hybrid cells undergo a reduction division, thus converting the hyperploid cell to a diploid karyotype and thereby concealing the fusion history.61 However, detection of donor-derived, diploid XY-chromosome–positive cells in recipient solid-organ tissue as early as 9 days15 and 13 days38 after allogeneic cell infusion makes this explanation unlikely. It has also been discussed whether hyperploid hepatocytes may provide a favorable environment for hybrid cells and may not be representative of other solid-organ tissues.65
Thus, cell fusion appears to account for the presence of cells in solid-organ tissue that show donor characteristics to some extent, but not completely. Fusion could even be seen as a physiologic ongoing repair mechanism by which cells deliver healthy and new genes to highly specialized cells to prevent them from dying and to correct genetically defective cells.66 Distinguishing between stem-cell fate transition and cell fusion demands that researchers use rigorous criteria in studying the phenomenon of stem-cell plasticity.
|
MODELS OF DIFFERENTIATION OF ADULT STEM CELLS INTO SOLID-ORGAN–SPECIFIC CELLS |
The results of transplantation studies involving sex-mismatched peripheral-blood stem cells suggest that the cells responsible for generating solid-organ–specific cells are, like hematopoietic stem cells, a group of circulating mononuclear cells. As outlined by Frisen,67 these studies of peripheral-blood stem-cell transplantation suggest four possible explanations for how adult stem cells derived from bone marrow or peripheral blood differentiate into nonlymphohematopoietic tissue cells (Figure 2). The first explanation is that multiple distinct types of stem cells circulate, with each type differentiating into its own lineage-restricted tissue. This deterministic model is supported by the fact that various progenitor cells with clonogenic potential circulate in the peripheral blood, including hematopoietic stem cells, mesenchymal stem cells,68 endothelial precursor cells,36,37,69 skeletal stem cells,70 and smooth-muscle progenitor cells.71,72 Therefore, it is conceivable that nonlymphohematopoietic, organ-specific stem cells, like hematopoietic stem cells, move between their own solid tissue and the peripheral blood.
|
|
|
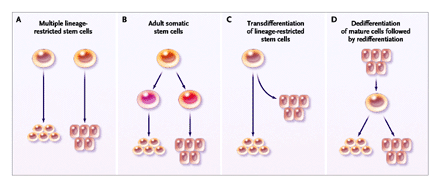
Figure 2. Various Models for Generating Solid-Organ Tissue Cells through Differentiation of Bone Marrow–Derived and Circulating Adult Stem Cells. |
[1] In the first model, distinct stem cells differentiate, each into its own organ-specific cell (Panel A). In the second model, primitive somatic stem cells located in hematopoietic tissue differentiate into various organ-specific cells (Panel B). In the third model, stem cells, such as hematopoietic stem cells, differentiate along their predetermined pathway. Under certain, probably rare conditions, tissue injury or another stimulus causes some stem cells to deviate from their predetermined pathway and generate cells of a different tissue — a process known as transdifferentiation (Panel C). In the fourth model, mature cells dedifferentiate into cells with stem-cell–like characteristics and eventually redifferentiate into terminally differentiated cells of their own tissue or a different tissue (Panel D).
[2] The second explanation postulates that a primordial equivalent to the embryonic stem cell, located in the bone marrow, peripheral blood, or both and available throughout adulthood, gives rise to various circulating, lineage-restricted stem cells. Evidence of such an adult somatic stem cell has been provided recently by Jiang et al.,73 who isolated multipotent adult progenitor cells from bone marrow–derived cultured cells; the progenitor cells were negative for CD34, CD44, CD45, c-kit, and major-histocompatibility-complex (MHC) classes I and II. These progenitor cells maintain their functional capacities when cultured for more than 100 population doublings, without obvious senescence, and differentiate in vivo into the hematopoietic lineage and into epithelium of the liver, lung, and gut. However, function of these end-differentiated cells within specific solid organs has not been proved.
[3] According to the third hypothesis, a circulating hematopoietic stem cell that expresses a default pathway of hematopoietic differentiation deviates, under certain conditions, from its preprogrammed pathway of differentiation and jumps the lineage barrier (a phenomenon known as transdifferentiation). Preclinical data support this hypothesis.18,19,20
[4] The fourth explanation is that a differentiated, solid-organ–specific cell regains characteristics similar to those of a stem cell in order to generate, through peripheral blood, differentiated cells of another tissue specificity (dedifferentiation and redifferentiation).3,74,75 In vivo dedifferentiation of epidermal cells to islands of stem cells in response to recombinant human epidermal growth factor has been reported.76
None of these four models of stem-cell differentiation have yet been proved to explain the mechanism underlying developmental stem-cell plasticity. In our opinion, preclinical data favor the transdifferentiation model, although this remains controversial. More than one model may be valid.
|
POTENTIAL ROLE OF CIRCULATING ADULT STEM CELLS IN TISSUE REPAIR |
Potential Role of Circulating Adult Stem Cells in Tissue Repair
It is well known that circulating hematopoietic stem cells are critical for hematopoietic homeostasis.77 Similarly, one may hypothesize that circulating stem cells contribute to homeostasis in solid-organ tissue.
|
MECHANISMS OF STEM-CELL RECRUITMENT AND DIFFERENTIATION |
The mechanisms by which circulating stem cells are recruited into various solid-organ tissues and tissue-specific cells are subsequently generated are not fully understood. Tissue injury that causes changes in the microenvironment may play an important part in stem-cell recruitment (Figure 3). In this regard, the highest percentage of donor-derived hepatocytes (about 40 percent) was found in a liver-transplant recipient with fibrosing cholestatic recurrent hepatitis C.28 A low frequency of hepatocyte replacement by bone marrow–derived stem cells has also been reported (up to 2.2 percent of total hepatocytes), even in the absence of histologic evidence of acute hepatic tissue injury,78 suggesting that circulating cells take part in tissue homeostasis. Because the model for most experimental and clinical transdifferentiation studies is allogeneic stem-cell transplantation with use of the Y chromosome as a marker, human leukocyte antigen disparity cannot be ruled out as a trigger for circulating stem-cell transdifferentiation.
|
|
|
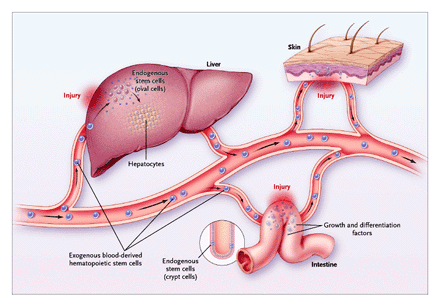
Figure 3. Possible Roles of Bone Marrow–Derived and Circulating Stem Cells in the Repair of Solid-Organ Tissue. |
After tissue injury, stem cells that are intrinsic to the tissue replace necrotic cells as a first line of defense. If the pool of endogenous stem cells is exhausted, exogenous circulating stem cells are signaled to replenish the pool and participate in tissue repair. Thus, circulating stem cells may serve as a backup rescue system.
A concentration of stem cells at the site of tissue damage has been shown to contribute to tissue repair. As noted earlier, Orlic and colleagues reported the repair of infarcted cardiac tissue after either the injection of stem cells derived from bone marrow directly into heart tissue surrounding the infarct15 or a systemic increase in the number of peripheral-blood hematopoietic stem cells to a level approximately 250 times as high as the base-line level with use of a combined treatment of recombinant human granulocyte colony-stimulating factor and stem-cell factor.25,79
The release of chemokines such as stromal-derived factor from injured hepatic tissue and interaction with its specific receptor, CXCR4, on hematopoietic precursor cells is another prototype of how tissue repair may function by aiding in the recruitment of circulating hematopoietic precursor cells to the site of tissue injury.80
On a molecular level, transcriptional regulation of hematopoietic stem cells may be altered in response to signals from the local environment. Specifically, homing, growth, and differentiation factors that may be involved in transitions of cell fate have been described for neuronal cells as a prototype.3
|
A CLINICAL MODEL OF SOLID-ORGAN TISSUE GENERATED BY CIRCULATING STEM CELLS |
Anderson and colleagues11 hypothesized that under physiologic steady-state conditions, an equilibrium is maintained whereby endogenous stem cells intrinsic to a specific tissue and, to a much lesser extent, blood-derived stem cells of heterologous lineage replenish apoptotic tissue. However, a rigorous demand for tissue repair that cannot be met by endogenous stem-cell differentiation may, as a backup system, trigger circulating stem cells to undergo differentiation into solid-organ tissue, though this is believed to be rare under physiologic steady-state conditions. Thus, the pool of tissue-intrinsic stem cells can be considered an intermediary between systemically circulating stem cells and terminally differentiated solid-organ–specific cells (Figure 3). Experimental evidence of a stepwise developmental progression from adult bone marrow through a stem cell intrinsic to muscle tissue to an adult muscle fiber in response to injury was provided by LaBarge and Blau.63 Adult cells derived from bone marrow contributed to the formation of up to 3.5 percent of differentiated muscle fibers. The triggering mechanisms for the plasticity of bone marrow-derived stem cells seemed to be an insufficient pool of stem cells intrinsic to this tissue in concert with an increased demand for the production of new cells and changes in the microenvironment as a result of tissue injury at the site.
|
POTENTIAL CLINICAL APPLICATIONS |
There are essentially two strategies for using adult stem cells derived from hematopoietic tissue for tissue repair. One approach is based on identifying and expanding in vitro multipotent adult progenitor cells that, like embryonic stem cells, are capable of generating mesodermal, ectodermal, and endodermal tissue.73 The other approach is based on the in vivo availability of a pool of systemic and circulating adult stem cells that can be manipulated to generate or repair solid-organ tissue.
Assuming that circulating stem cells generate cells specific to solid organs in vivo, a potential clinical concept of tissue repair would require three conditions. First, the stem-cell pool must be easily accessible, as is the case with the circulating stem-cell pool that is routinely used for harvesting hematopoietic stem cells.81 Second, the concentration of stem cells at the site of tissue regeneration must be sufficient, which can be accomplished either by cytokine-induced mobilization of hematopoietic stem cells,81 mesenchymal stem cells,59 angioblasts,36,69 and smooth-muscle progenitor cells71,72 from extravascular sites into the circulating blood or by directly delivering the cells to the site of tissue injury. Third, appropriate signals from the site of damaged tissue must direct exogenous stem cells to the site where they are needed. However, we do not yet clearly understand how to manipulate the microenvironment surrounding the area of tissue regeneration actively and signal exogenous blood-derived stem cells to participate in tissue regeneration in vivo.
As with transplantation of hematopoietic stem cells, solid-organ tissue repair can be accomplished by differentiation of autologous or allogeneic stem cells. Allogeneic transplants are advantageous in experimental settings in which the Y chromosome is used as a marker, as well as in hereditary stem-cell disorders, in which transplantation of normal stem cells is supposed to generate normally functioning tissue. Differentiation of autologous bone marrow or circulating stem cells into solid-organ tissue seems the preferred therapeutic approach, as it precludes immunologic disparities.
It should be emphasized that the interpretation of some of the clinical data reported so far is controversial, and in some cases there is no confirmation of their validity. Selected potential clinical indications for generating solid-organ tissue with the use of stem cells derived from hematopoietic tissue are listed in Table 2.
|
EMBRYONIC STEM CELLS |
Multipotent adult progenitor cells are similar to embryonic stem cells in that they can be extensively expanded in vivo, and have ‘extensive proliferation and clonal multilineage differentiation potential’73 in vitro and in vivo to form cells of three germ layers. In contrast to embryonic stem cells, multipotent adult progenitor cells can be harvested from autologous bone marrow, thus avoiding the rejection that results from the expression of MHC proteins in human embryonic stem cells.82 Immunologic disparities associated with the use of embryonic stem cells may be overcome by applying the technique of somatic-cell nuclear transfer (therapeutic cloning). A major concern with the use of embryonic stem cells for therapeutic purposes is their potential to form teratomas in vivo; such tumorigenicity is thus far unknown for pluripotent adult progenitor cells.83,84 The in vivo differentiation potential of circulating adult stem cells into nonlymphohematopoietic tissue under steady-state conditions seems to be inferior to that of ex vivo expanded embryonic stem cells and multipotent adult progenitor cells. However, the extent to which in vivo manipulation of circulating adult stem cells and their target tissue can induce terminal cell differentiation for repairing or replacing solid-organ tissue remains to be demonstrated.
|
CONCLUSIONS AND OUTLOOK |
We are only beginning to understand the circulating blood, not only as a distributor of hematopoietic progenitor cells but also as a systemic supplier of progenitor cells that have the potential to participate in the homeostasis of various solid-organ tissues and that are capable of nonlymphohematopoietic tissue repair. There is a growing body of evidence that the adult stem-cell system may be more flexible than previously thought, particularly under conditions of tissue stress. As a consequence, tissue-specific cell systems may be regarded as versatile and dynamic, irrespective of lineage-specific restrictions.
The observation that solid-organ tissues are colonized by organ-specific cells originating from the circulating blood suggests that tissue regeneration and repair may be feasible if we learn to direct progenitor cells from the circulating blood into areas of injured or diseased tissue and to modulate their proliferation and maturation once these cells have reached the target tissue. Despite the promising studies indicating the potential plasticity of adult stem cells, many obstacles remain, some of which may favor the misinterpretation of data that indicate such a phenomenon. The basic mechanisms of stem-cell differentiation that lead to the formation of solid-organ tissue are still not completely understood. However, translational research, including clinical studies, is already being performed to develop potential treatment strategies.
We are indebted to Kate O Suilleabhain and David Galloway for their editorial assistance.
SOURCE INFORMATION
From the Department of Blood and Marrow Transplantation (M.K.) and the Department of Bioimmunotherapy (Z.E.), the University of Texas M.D. Anderson Cancer Center, Houston.
Address reprint requests to Dr. Körbling at the University of Texas M.D. Anderson Cancer Center, Department of Blood and Marrow Transplantation, Unit 423, 1515 Holcombe Blvd., Houston, TX 77030, or at mkorblin@mdanderson.org.
References
1. Slack JMW. Stem cells in epithelial tissues. Science 2000;287:1431-1433.[Abstract/Full Text]
2. Weissman IL. Stem cells: units of development, units of regeneration, and units in evolution. Cell 2000;100:157-168.[ISI][Medline]
3. Blau HM, Brazelton TR, Weimann JM. The evolving concept of a stem cell: entity or function? Cell 2001;105:829-841.[CrossRef][ISI][Medline]
4. Quesenberry PJ, Colvin GA, Lambert J-F. The chiaroscuro stem cell: a unified stem cell theory. Blood 2002;100:4266-4271.[Abstract/Full Text]
5. Graf T. Differentiation plasticity of hematopoietic cells. Blood 2002;99:3089-3101.[Full Text]
6. Ferrari G, Cusella-De Angelis G, Coletta M, et al. Muscle regeneration by bone marrow-derived myogenic progenitors. Science 1998;279:1528-1530.[Abstract/Full Text]
7. Petersen BE, Bowen WC, Patrene KD, et al. Bone marrow as a potential source of hepatic oval cells. Science 1999;284:1168-1170.[Abstract/Full Text]
8. Eglitis MA, Mezey E. Hematopoietic cells differentiate into both microglia and macroglia in the brains of adult mice. Proc Natl Acad Sci U S A 1997;94:4080-4085.[Abstract/Full Text]
9. Mezey E, Chandross KJ, Harta G, Maki RA, McKercher SR. Turning blood into brain: cells bearing neuronal antigens generated in vivo from bone marrow. Science 2000;290:1779-1782.[Abstract/Full Text]
10. Brazelton T, Rossi FM, Keshet GI, Blau HM. From marrow to brain: expression of neuronal phenotypes in adult mice. Science 2000;290:1775-1779.[Abstract/Full Text]
11. Anderson DJ, Gage FH, Weissman IL. Can stem cells cross lineage boundaries? Nat Med 2001;7:393-395.[CrossRef][ISI][Medline]
12. Gussoni E, Soneoka Y, Strickland CD, et al. Dystrophin expression in the mdx mouse restored by stem cell transplantation. Nature 1999;401:390-394.[CrossRef][ISI][Medline]
13. Lagasse E, Connors H, Al-Dhalimy M, et al. Purified hematopoietic stem cells can differentiate into hepatocytes in vivo. Nat Med 2000;6:1229-1234.[CrossRef][ISI][Medline]
14. Wang X, Ge S, McNamara G, Hao Q-L, Crooks GM, Nolta JA. Albumin-expressing hepatocyte-like cells develop in the livers of immune-deficient mice that received transplants of highly purified human hematopoietic stem cells. Blood 2003;101:4201-4208.[Abstract/Full Text]
15. Orlic D, Kajstura J, Chimenti S, et al. Bone marrow cells regenerate infarcted myocardium. Nature 2001;410:701-705.[CrossRef][ISI][Medline]
16. Jackson KA, Majka SM, Wang H, et al. Regeneration of ischemic cardiac muscle and vascular endothelium by adult stem cells. J Clin Invest 2001;107:1395-1402.[Abstract/Full Text]
17. Sata M, Saiura A, Kunisato A, et al. Hematopoietic stem cells differentiate into vascular cells that participate in the pathogenesis of arteriosclerosis. Nat Med 2002;8:403-409.[CrossRef][ISI][Medline]
18. Krause DS, Theise ND, Collector MI, et al. Multi-organ, multi-lineage engraftment by a single bone marrow-derived stem cell. Cell 2001;105:369-377.[ISI][Medline]
19. Grant MB, May WS, Caballero S, et al. Adult hematopoietic stem cells provide functional hemangioblast activity during retinal neovascularization. Nat Med 2002;8:607-612.[CrossRef][ISI][Medline]
20. Masuya M, Drake CJ, Fleming PA, et al. Hematopoietic origin of glomerular mesangial cells. Blood 2003;101:2215-2218.[Abstract/Full Text]
21. Eschbach JW Jr, Epstein RB, Burnell JM, Thomas ED. Physiologic observations in human cross circulation. N Engl J Med 1965;273:997-1003.[Medline]
22. Wright DE, Wagers AJ, Gulati AP, Johnson FL, Weissman IL. Physiological migration of hematopoietic stem and progenitor cells. Science 2001;294:1933-1936.[Abstract/Full Text]
23. Takahashi T, Kalka C, Masuda H, et al. Ischemia- and cytokine-induced mobilization of bone marrow-derived endothelial progenitor cells for neovascularization. Nat Med 1999;5:434-438.[CrossRef][ISI][Medline]
24. Kocher AA, Schuster MD, Szabolcs MJ, et al. Neovascularization of ischemic myocardium by human bone-marrow-derived angioblasts prevents cardiomyocyte apoptosis, reduces remodeling and improves cardiac function. Nat Med 2001;7:430-436.[CrossRef][ISI][Medline]
25. Orlic D, Kajstura J, Chimenti S, et al. Mobilized bone marrow cells repair the infarcted heart, improving function and survival. Proc Natl Acad Sci U S A 2001;98:10344-10349.[Abstract/Full Text]
26. Orlic D, Arai AE, Sheikh FH, et al. Cytokine mobilized CD34+ cells do not benefit rhesus monkeys following induced myocardial infarction. Blood 2002;100:Suppl 1:28a-29a. abstract.
27. Horwitz EM, Prockop DJ, Fitzpatrick LA, et al. Transplantability and therapeutic effects of bone marrow-derived mesenchymal cells in children with osteogenesis imperfecta. Nat Med 1999;5:309-313.[CrossRef][ISI][Medline]
28. Theise ND, Nimmakayalu M, Gardner R, et al. Liver from bone marrow in humans. Hepatology 2000;32:11-16.[CrossRef][ISI][Medline]
29. Alison MR, Poulsom R, Jeffery R, et al. Hepatocytes from non-hepatic adult stem cells. Nature 2000;406:257-257.[CrossRef][ISI][Medline]
30. Kleeberger W, Rothamel T, Glockner S, Flemming P, Lehmann U, Kreipe H. High frequency of epithelial chimerism in liver transplants demonstrated by microdissection and STR-analysis. Hepatology 2002;35:110-116.[CrossRef][ISI][Medline]
31. Okamoto R, Yajima T, Yamazaki M, et al. Damaged epithelia regenerated by bone marrow-derived cells in the human gastrointestinal tract. Nat Med 2002;8:1011-1017.[CrossRef][ISI][Medline]
32. Mezey E, Key S, Vogelsang G, Szalayova I, Lange GD, Crain B. Transplanted bone marrow generates new neurons in human brains. Proc Natl Acad Sci U S A 2003;100:1364-1369.[Abstract/Full Text]
33. Weimann JM, Charlton GA, Brazelton TR, Hackman RC, Blau H. Contribution of transplanted bone marrow cells to Purkinje neurons in human adult brains. Proc Natl Acad Sci U S A 2003;100:2088-2093.[Abstract/Full Text]
34. Quaini F, Urbanek K, Beltrami A, et al. Chimerism of the transplanted heart. N Engl J Med 2002;346:5-15.[Abstract/Full Text]
35. Müller P, Pfeiffer P, Koglin J, et al. Cardiomyocytes of noncardiac origin in myocardial biopsies of human transplanted hearts. Circulation 2002;106:31-35.[Abstract/Full Text]
36. Lagaaij EL, Cramer-Knijnenburg GF, van Kemenade FJ, van Es LA, Bruijn JA, van Krieken JH. Endothelial cell chimerism after renal transplantation and vascular rejection. Lancet 2001;357:33-37.[CrossRef][ISI][Medline]
37. Lin Y, Weisdorf DJ, Solovey A, Hebbel RP. Origins of circulating endothelial cells and endothelial outgrowth from blood. J Clin Invest 2000;105:71-77.[Abstract/Full Text]
38. Körbling M, Katz RL, Khanna A, et al. Hepatocytes and epithelial cells of donor origin in recipients of peripheral-blood stem cells. N Engl J Med 2002;346:738-746.[Abstract/Full Text]
39. Hematti P, Sloand EM, Carvallo CA, et al. Absence of donor-derived keratinocyte stem cells in skin tissue cultured from patients after mobilized peripheral blood hematopoietic stem cell transplantation. Exp Hematol 2002;30:943-949.[CrossRef][ISI][Medline]
40. Tran SD, Pillemer SR, Dutra A, et al. Differentiation of human bone marrow-derived cells into buccal epithelial cells in vivo: a molecular analytical study. Lancet 2003;361:1084-1088.[CrossRef][ISI][Medline]
41. Eto T, Takahashi H. Enhanced inhibition of hepatitis B virus production by asialo-glycoprotein receptor-directed interferon. Nat Med 1999;5:577-581.[CrossRef][ISI][Medline]
42. Ueki T, Kaneda Y, Tsutsui H, et al. Hepatocyte growth factor gene therapy of liver cirrhosis in rats. Nat Med 1999;5:226-230.[CrossRef][ISI][Medline]
43. Strauer BE, Brehm M, Zeus T, et al. Repair of infarcted myocardium by autologous intracoronary mononuclear bone marrow cell transplantation in humans. Circulation 2002;106:1913-1918.[Abstract/Full Text]
44. Stamm C, Westphal B, Kleine H-D, et al. Autologous bone-marrow stem-cell transplantation for myocardial regeneration. Lancet 2003;361:45-46.[CrossRef][ISI][Medline]
45. Assmus B, Schachinger V, Teupe C, et al. Transplantation of Progenitor Cells and Regeneration Enhancement in Acute Myocardial Infarction (TOPCARE-AMI). Circulation 2002;106:3009-3017.[Abstract/Full Text]
46. Tse H-F, Kwong Y-L, Chan JKF, Lo G, Ho C-L, Lau CP. Angiogenesis in ischaemic myocardium by intramyocardial autologous bone marrow mononuclear cell implantation. Lancet 2003;361:47-49.[CrossRef][ISI][Medline]
47. Perin EC, Dohmann HFR, Borojevic R, et al. Transendocardial, autologous bone marrow cell transplantation for severe, chronic ischemic heart failure. Circulation 2003;107:2294-2302.[Abstract/Full Text]
48. Edelberg JM, Tang L, Hattori K, Lyden D, Rafii S. Young adult bone marrow-derived endothelial precursor cells restore aging-impaired cardiac angiogenic function. Circ Res 2002;90:E89-E93.[CrossRef][ISI][Medline]
49. Tateishi-Yuyama E, Matsubara H, Murohara T, et al. Therapeutic angiogenesis for patients with limb ischaemia by autologous transplantation of bone-marrow cells: a pilot study and a randomised controlled trial. Lancet 2002;360:427-435.[CrossRef][ISI][Medline]
50. Otani A, Kinder K, Ewalt K, Otero FJ, Schimmel P, Friedlander M. Bone marrow-derived stem cells target retinal astrocytes and can promote or inhibit retinal angiogenesis. Nat Med 2002;8:1004-1010.[CrossRef][ISI][Medline]
51. Gussoni E, Bennett RR, Muskiewicz KR, et al. Long-term persistence of donor nuclei in a Duchenne muscular dystrophy patient receiving bone marrow transplantation. J Clin Invest 2002;110:807-814.[Abstract/Full Text]
52. Chen J, Sanberg PR, Li Y, et al. Intravenous administration of human umbilical cord blood reduces behavioral deficits after stroke in rats. Stroke 2001;32:2682-2688.[Abstract/Full Text]
53. Taylor DA, Hruban R, Rodriguez R, Goldschmidt-Clermont PJ. Cardiac chimerism as a mechanism for self-repair: does it happen and if so to what degree? Circulation 2002;106:2-4.[Full Text]
54. Castro RF, Jackson KA, Goodell MA, Robertson CS, Liu H, Shine HD. Failure of bone marrow cells to transdifferentiate into neural cells in vivo. Science 2002;297:1299-1299.[Full Text]
55. Wagers AJ, Sherwood RI, Christensen JL, Weissman IL. Little evidence for developmental plasticity of adult hematopoietic stem cells. Science 2002;297:2256-2259.[Abstract/Full Text]
56. Blau H, Brazelton T, Keshet G, Rossi F. Something in the eye of the beholder. Science 2002;298:361-363.
57. Mezey E, Nagy A, Szalayova I, et al. Comment on: ‘Failure of bone marrow cells to transdifferentiate into neural cells in vivo.’ Science 2003;299:1184-1184.[ISI]
58. Theise ND, Krause DS, Sharkis S. Comment on: ‘Little evidence for developmental plasticity of adult hematopoietic stem cells.’ Science 2003;299:1317-1317.[Medline]
59. Terada N, Hamazaki T, Oka M, et al. Bone marrow cells adopt the phenotype of other cells by spontaneous cell fusion. Nature 2002;416:542-545.[CrossRef][ISI][Medline]
60. Ying QL, Nichols J, Evans E, Smith AG. Changing potency by spontaneous fusion. Nature 2002;416:545-548.[CrossRef][ISI][Medline]
61. Wang X, Willenbring H, Akkari Y, et al. Cell fusion is the principal source of bone-marrow-derived hepatocytes. Nature 2003;422:897-901.[CrossRef][ISI][Medline]
62. Vassilopoulos G, Wang PR, Russell DW. Transplanted bone marrow regenerates liver by cell fusion. Nature 2003;422:901-904.[CrossRef][ISI][Medline]
63. LaBarge MA, Blau HM. Biological progression from adult bone marrow to mononucleate muscle stem cell to multinucleate muscle fiber in response to injury. Cell 2002;111:589-601.[ISI][Medline]
64. Srivatsa B, Srivatsa S, Johnson KL, Samura O, Lee SL, Bianchi DW. Microchimerism of presumed fetal origin in thyroid specimens from women: a case-control study. Lancet 2001;358:2034-2038.[CrossRef][ISI][Medline]
65. Medvinsky A, Smith A. Fusion brings down barriers. Nature 2003;422:823-825.[CrossRef][ISI][Medline]
66. Blau HM. A twist of fate. Nature 2002;419:437-437.[CrossRef][ISI][Medline]
67. Frisen J. Stem cell plasticity? Neuron 2002;35:415-418.[ISI][Medline]
68. Huss R, Lange C, Weissinger EM, Kolb HJ, Thalmeier K. Evidence of peripheral blood-derived, plastic-adherent CD34-/low hematopoietic stem cell clones with mesenchymal stem cell characteristics. Stem Cells 2000;18:252-260.[Abstract/Full Text]
69. Masuda H, Kalka C, Asahara T. Endothelial progenitor cells for regeneration. Hum Cell 2000;13:153-160.[Medline]
70. Kuznetsov SA, Mankani MH, Gronthos S, Satomura K, Bianco P, Robey PG. Circulating skeletal stem cells. J Cell Biol 2001;153:1133-1139.[Abstract/Full Text]
71. Saiura A, Sata M, Hirata Y, Nagai R, Makuuchi M. Circulating smooth muscle progenitor cells contribute to atherosclerosis. Nat Med 2001;7:382-383.[CrossRef][ISI][Medline]
72. Simper D, Stalboerger PG, Panetta CJ, Wang S, Caplice NM. Smooth muscle progenitor cells in human blood. Circulation 2002;106:1199-1204.[Abstract/Full Text]
73. Jiang Y, Jahagirdar BN, Reinhardt RL, et al. Pluripotency of mesenchymal stem cells derived from adult marrow. Nature 2002;418:41-49.[CrossRef][ISI][Medline]
74. Odelberg SJ, Kollhoff A, Keating MT. Dedifferentiation of mammalian myotubes induced by msx1. Cell 2000;103:1099-1109.[ISI][Medline]
75. Kondo T, Raff M. Oligodendrocyte precursor cells reprogrammed to become multipotential CNS stem cells. Science 2000;289:1754-1757.[Abstract/Full Text]
76. Fu X, Sun X, Li X, Sheng Z. Dedifferentiation of epidermal cells to stem cells in vivo. Lancet 2001;358:1067-1068.[CrossRef][ISI][Medline]
77. Wright DE, Bowman EP, Wagers AJ, Butcher EC, Weissman IL. Hematopoietic stem cells are uniquely selective in their migratory response to chemokines. J Exp Med 2002;195:1145-1154.[Abstract/Full Text]
78. Theise ND, Badve S, Saxena R, et al. Derivation of hepatocytes from bone marrow cells in mice after radiation-induced myeloablation. Hepatology 2000;31:235-240.[ISI][Medline]
79. Bodine DM, Seidel NE, Gale MS, Nienhuis AW, Orlic D. Efficient retrovirus transduction of mouse pluripotent hematopoietic stem cells mobilized into the peripheral blood by treatment with granulocyte colony-stimulating factor and stem cell factor. Blood 1994;84:1482-1491.[Abstract/Full Text]
80. Hatch HM, Zheng D, Jorgensen ML, Petersen BE. SDF-1/CXCR4: a mechanism for hepatic oval cell activation and bone marrow stem cell recruitment to the injured liver of rats. Cloning Stem Cells 2002;4:339-51.
81. Körbling M, Anderlini P. Peripheral blood stem cell versus bone marrow allotransplantation: does the source of hematopoietic stem cells matter? Blood 2001;98:2900-2908.[Abstract/Full Text]
82. Drukker M, Katz G, Urbach A, et al. Characterization of the expression of MHC proteins in human embryonic stem cells. Proc Natl Acad Sci U S A 2002;99:9864-9869.[Abstract/Full Text]
83. Odorico JS, Kaufman DS, Thomson JA. Multilineage differentiation from human embryonic stem cell lines. Stem Cells 2001;19:193-204.[Abstract/Full Text]
84. Grompe M. Adult versus embryonic stem cells: it's still a tie. Mol Ther 2002;6:303-305.[CrossRef][ISI][Medline]
Table 1. Adult Human Stem Cells and Their Primary Direction of Differentiation.
|
Cell Type |
Tissue-Specific Location |
Cells or Tissues Produced |
|
Hematopoietic |
Bone marrow, peripheral blood |
Bone marrow and blood lymphohematopoetic cells |
|
Mesenchymal stem cell |
Bone marrow, peripheral blood |
Bone, cartilage, tendon, adipose tissue, muscle, marrow stroma, neural cells |
|
Neural stem cells |
Ependymal cells, astrocytes (subventricular zone) of the CNS |
Neurons, astrocytes, oligodendrocytes |
|
Hepatic stem cells |
In or near the terminal bile ductules (canals of Hering) |
Oval cells that subsequently generate hepatocytes and ductular cells |
|
Pancreatic Stem cells |
Intra-islet Nestin positive cells, oval cells, duct cells |
Beta cells |
|
Skeletal muscle Stem cells |
Muscle fibers |
Skeletal muscle fibers |
|
Stem cells of the skin (keratinocytes) |
Basal layer of the epidermis, bulge zone of the hair follicles |
Epidermis, hair follicles |
|
Epithelial stem cells of the lung |
Tracheal basal and mucus-secreting cells, bronciolar Clara cells, alveolar Type II pneumocyte |
Mucous and ciliated cells, type I and II pneumocytes |
|
Stem cells of the intestinal epithelium |
Epithelial cells located around the base of each crypt |
Paneth’s cells, brush-border enterocytes, mucus secreting goblet cells, enteroendocrine cells |